
Extending our Wine Flavor Chemistry infographic to Whisky, we found there was alot of chemical overlap between flavor molecules found in whisky and the fruits and spices that whisky can taste like!

Extending our Wine Flavor Chemistry infographic to Whisky, we found there was alot of chemical overlap between flavor molecules found in whisky and the fruits and spices that whisky can taste like!
Comments Off on Whisky Flavor Chemistry
Posted in Uncategorized

This is a map of academic cancer-research in the United States organized by university. The area of each university label is proportional to the number of cancer-related papers published in the past 5 years (as downloaded from PubMed).
In addition, collaborations between universities are represented as flight edges where the thickness & transparency of each edge is proportional the the number of published collaborations between universities within that same time period.
Comments Off on Cancer Research in the USA
Posted in Uncategorized
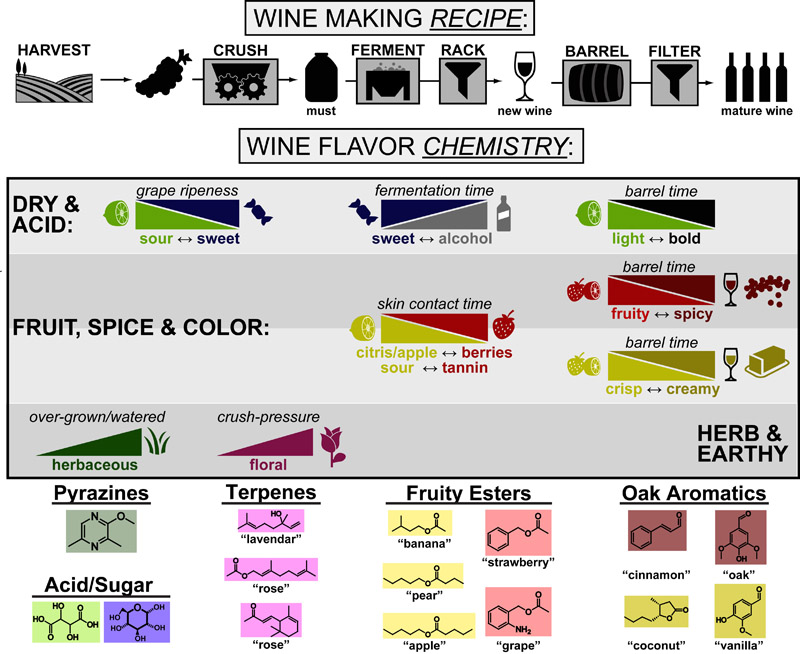
Our recent whiskey-graphic, led to some requests for a similar graphic on wine. Based on our research, wine appears to have a simpler “process chemistry” but more complicated “flavor chemistry” than whiskey. While we are still anchoring this graphic in the “production process chemistry” its major focus is on how individual steps affect the flavor of a wine.

A recent trip along the Bourbon Trail of Kentucky inspired the above summary (which started as rough notes from tours of four distilleries) which compares: (1) Whiskey Production Recipe (2) Whiskey Production Chemistry and (3) Distinguishing characteristics of major whiskey types. More detailed information can be found in the references below.
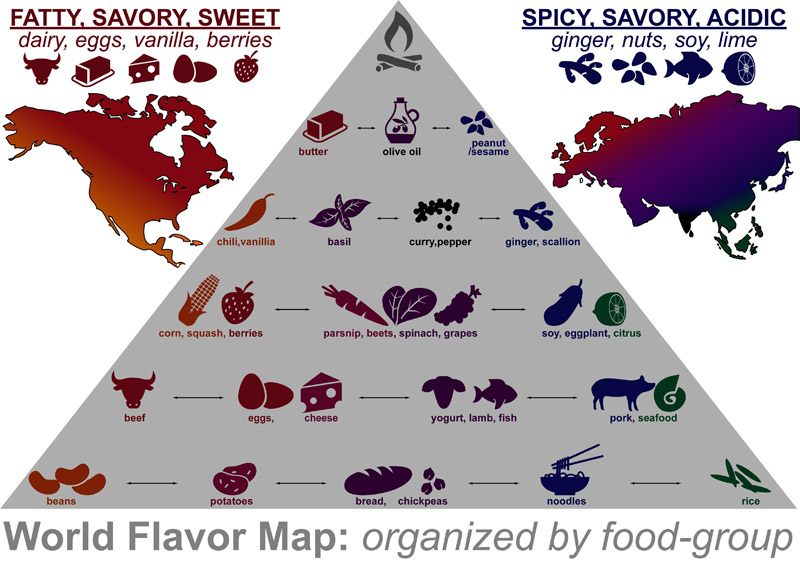
The graphic above summarizes ingredients that are most characteristic of a world-region’s recipes. It was inspired by a 2011 paper that examined connections between RECIPES – INGREDIENTS – FLAVOR-CHEMICALS.1 A major finding of this paper was that: A handful of common ingredients best distinguish and define Western and Eastern recipes [and flavor-chemical networks].
The graphic below provides a more detailed description on region-specific ingredients:
Comments Off on A World Flavors Map
Posted in Cooking
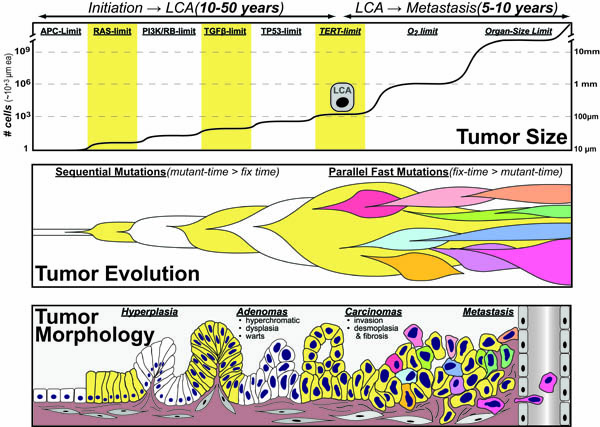
Tumors develop in two distinct stages: (1) Initiation to Last-Common-Ancestor(LCA) where 6-8 “driver-mutations” occur sequentially over 10-50 years and (2) LCA to Metastasis where mutations occur more rapidly in parallel. In the first phase, mutations occur slowly and pierce 6-8 barriers to unrestrained growth by “hot-wiring” pro-growth pathways and then “shorting-out” pro-death pathways . In the second phase, genomic instability creates a large-heterogeneous-population that is well-equipped to compete against bigger-picture-limitations such as oxygen-availability, tissue-size, attacking immune cells and chemotherapies.
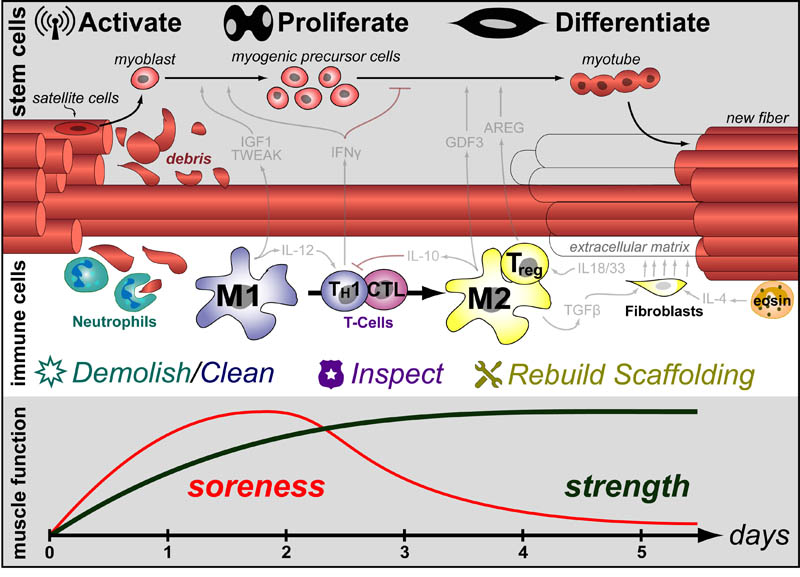
We’ve made several timelines on the adaptive-response that follows intense exercise ranging from: 2-3 months it takes to “get in shape” to the the 12 hours it takes to cement muscle-memory in the brain. One topic we have not discussed is the 1-week it takes to recover-and-rebuild from an intense bout of exercise.
Interesting, it is the immune-system (not stem cells) that manage the whole repair and rebuilding process. To use a building analogy, the immune system: (1) demolishes the old building (damaged muscle) and (2) erects the steel-sub-structure (extracellular matrix). Only then do the stem-cells install plumbing, electricity and internet (new muscle). On other words, the immune-cells “build the building” while the stem-cells “make it functional.”
Comments Off on Intense-Exercise Recovery Timeline (~1 week)
Posted in Running
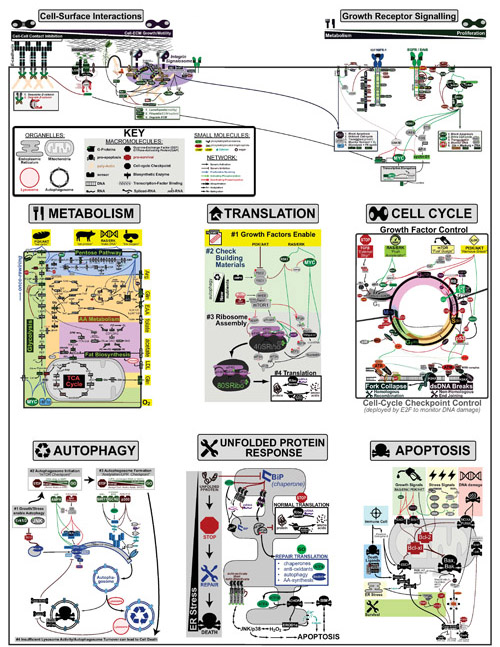
Cancer is a complex disease that is defined by at least 10 different “Hallmarks” that reflect mutation or epigenetically-driven reprogramming of normal cellular circuits. Over the past year, I have compiled a document that attempts to combine what’s known about the intra-cellular network that underlie these “Cancer Hallmarks.” This project started out as a single-page infographic but has since expanded into the 2 foot x 3 foot poster pictured above.
My goal was (and is) to create a comprehensive network map that is conceptually accessible to help me (and now others) think about the “big picture” of cancer networks. Of course, this poster is a work in progress and I will continue to update it over time. Below, I give a brief conceptual description of each module I have used to organize this “Canonical” Cancer-Network Map.
Posted in Systems Biology

I recently finished two excellent books (The Biology of Cancer and The Death of Cancer) that really helped me start to understand the “big picture” of cancer research (in the lab AND the clinic). These books inspired us to piece together a timeline on the history of cancer therapy which ended up being a bit larger than we expected. Click on the image above for a PDF of our Cancer Therapy Poster (3′ x 1′). Below you can find a summary of all the abbreviations and references used for this project.
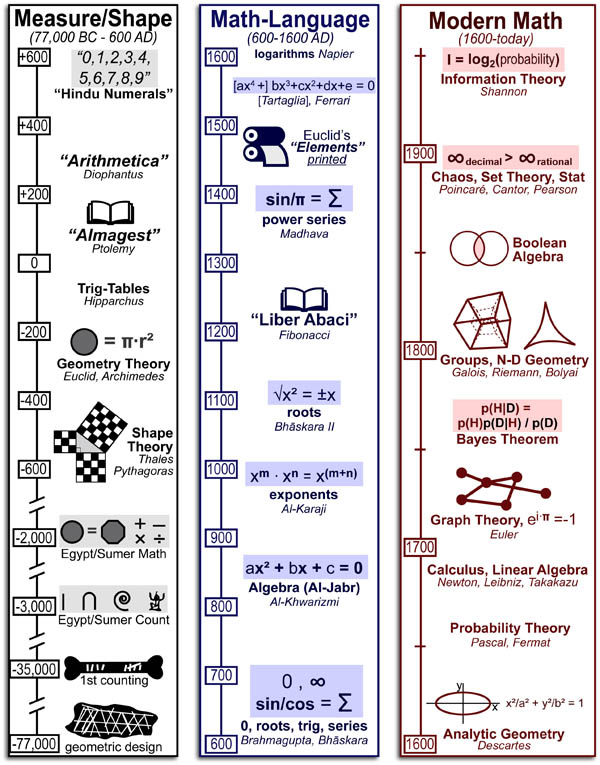
The history of mathematics can be divided into three periods:
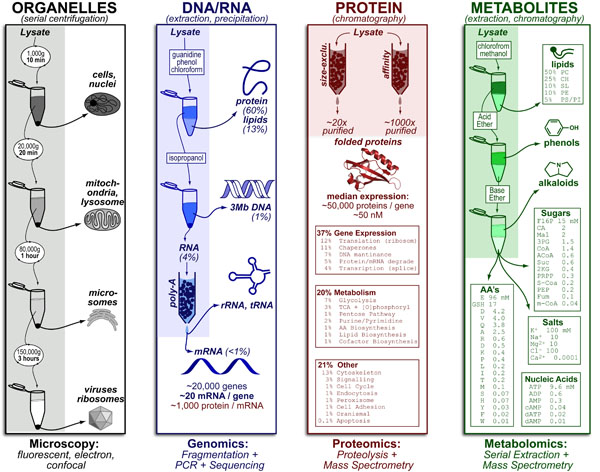
Different scientific questions focus on different parts of the cell and it is often necessary to break a cell up into those different pieces (figure above). While various “-omic” methods are well suited to answering global/systems-level questions for the four catagories listed above (e.g. microscopy, genomics, proteomics, metabolomics) they often lack the resolution of fractionation-methods to answer molecular level questions.
Comments Off on How do you “Fractionate” a Cell?
Posted in Cellular Biology

Quantitative methods in science can be categorized via their typical place within the scientific method as (1) Inferential which is focused primarily on data analysis and (2) Predictive which is focused on formulating mechanistic hypotheses through modeling. In the figure above we summarize some of the most common methods that fall within each of these categories.
Comments Off on Data-Inference vs Predictive-Modeling
Posted in Systems Biology
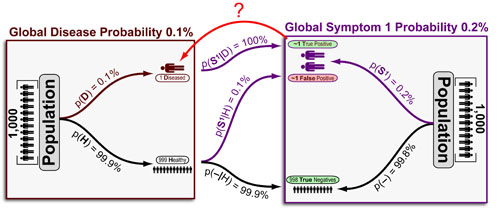
Baysian statistical inference is a very useful method to “back predict” the probability of a hypotheses from data frequency. In the example above, our “hypothesis” is “a disease“ and our “data” is the “an associated symptom.” Now, diseases are not measured directly, but rather, are diagnosed based on a combination of symptoms. Bayesian inference allows us to calculate the “Probability of Disease given Symptom 1 (p(D|S1)) with the following information:
Continue reading
Comments Off on Introduction to Bayesian Inference
Posted in Systems Biology
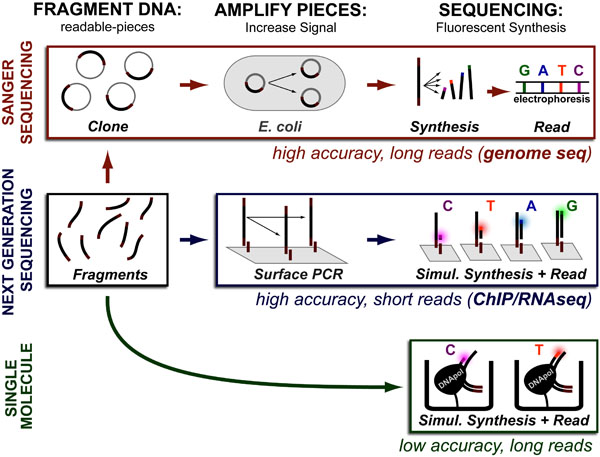
While DNA-sequencing methods are diverse and complex they can be grouped into three categories which share several common features: 1. DNA Fragmentation, 2. Fragment Amplification, 3. Sequencing via Fluorescent-Synthesis. These categories are:
Continue reading
Comments Off on DNA Sequencing Methods
Posted in Molecular Biology, Systems Biology
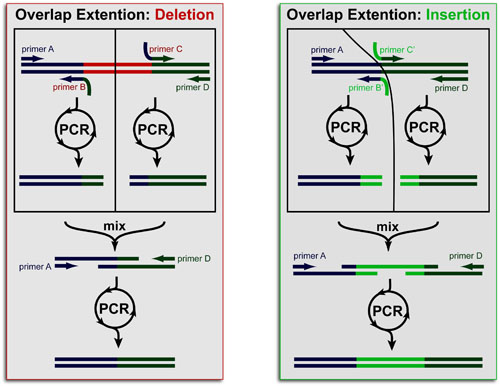
Polymerase Chain-Reaction (PCR) has become the backbone of most Methods in Molecular Biology and site-specific mutagenesis no exception. The key to PCR-based mutation of DNA is careful design of primers. In the simplest case, a point-mutation can be inserted into all PCR products by adding a point mutation to all primers. Unfortunately, for linear DNA, this method only works for mutagenesis at the ends of the template (where the primers bind).
Overlap extension, is a powerful 2-step, multi-PCR technique that can insert mutations at any position and of any size (including whole deletions or insertions). It accomplished this using chimeric primers to (1) cut out pieces of DNA and (2) reassemble them at overlap points:
Continue reading
Comments Off on PCR Mutagenesis: Overlap Extension
Posted in Molecular Biology